第二章、地中海世界的生命
时间: 2021-09-23 17:39:14 | 作者:清青水草 | 来源: 喜蛋文章网 | 编辑: admin | 阅读: 108次
- 2023-11-29 04:59:20麻老虎是什么生物
- 2023-11-24 12:00:58世界历史纷繁复杂,如何从中理出头绪
- 2023-11-14 21:01:30世界历史上,有哪些「急流勇退」的真实历史人物和真实历史事件
- 2023-11-02 10:00:10为什么电影《流浪地球2》中未来几乎没有生物方面的突破
- 2023-10-27 22:01:57地球上的生物,为什么只有人类进化出了“手”
- 2023-10-27 11:01:59为什么历史书不从分子生物学的角度讲人类迁移史
- 2023-10-11 09:00:36生物学小科普丨为什么地球生命的共同祖先名叫LUCA
- 2023-10-10 17:01:18世界上最高的人是瑞恩诺斯么 古生物学家估计他直立时肩膀处将近5米高是真的么
- 2023-10-10 17:01:11请问原始地球中生命现象如何诞生 有机生物是如何从无机物中诞生
- 2023-09-29 09:01:08世界历史为什么给人一种风水轮流转的感觉 为什么总是你方唱罢我登场,一个国家为什么不能永远的强盛下去

接下来,我们来谈谈地中海世界的生命。在讨论之前,我们需要明晰一些基本事实和概念。生命的本质是什么呢?科学家们认为,生命是一群能摄入各种高能物料,以保持自身存续和复制繁衍的分子聚合体。令人惊讶的是,所有生命(主要指当前三域系统内细菌域(Bacteria)、古菌域(Archaea)和真核域(Eukarya)的生命,一些特殊的生命如病毒等并不一定包括在内)都来自一个共同的祖先,这是因为这些生命都有共同的基本生化组织,比如说由氨基酸构成的蛋白质、由DNA或RNA序列构成的遗传密码、标准化的储能方式——三磷酸腺苷ATP等。以ATP为例,三磷酸腺苷分子中有高能的化学键,当高能化学键断裂时,键能就会转移到其他分子中,同时三磷酸腺苷变为相对低能的二磷酸腺苷ADP。如果要重新合成三磷酸腺苷分子,需要某些特殊的酶催化,将能量从能源分子转移到二磷酸腺苷中。三磷酸腺苷的能量会被转化成动能(人走动)、光能(萤火虫灯光)、电能(电鳗放电)等,但无论最终变成什么形式的能量,生物利用的所有能量本质上都是来自三磷酸腺苷的化学能。如果细胞想用能源物质比如葡萄糖来合成自身的成分,它会将葡萄糖里的化学能转移到三磷酸腺苷里,而后三磷酸腺苷的化学能被用于合成其他分子。我们知道,地球上的生命种类繁多,单独的共同祖先是如何分化出形态各异的子孙后代的?进化论解释了这个问题。生物的遗传信息储存在其基因上,生物将遗传信息传递给后代时,有极小概率产生偶然的基因复制错误,这些错误导致子代与上一代的些微不同,这种不同称之为基因变异。基因的组合和变异使得这些子代的性状(如人的身高、肤色、发色等)与上一代不同。在生存繁衍竞争中,适应环境的变异会在生物种群中留存下来,不适应环境的变异个体留下子代的概率降低,其变异基因也逐渐消失,这在进化论中称之为“自然选择”。生命是渐进的。从15亿年前的有核细胞,到6亿年前的脊椎动物,2.45亿年前的哺乳动物,到5500万年前的灵长类出现,到现在的我们,每个阶段都有所改变。不过,我们独特的属性,就像是一栋多房间大楼的加盖部分,其中大部分的房间与其他物种共享。比如,我们和线虫有相同的控制食欲的基因。我们是一栋由物种演化搭盖成的复式大楼。我们现在了解到,基因的变异空间及其巨大。生物拥有三大基因网络:1、代谢基因网络,生物利用生物化学反应,获取能量的控制基因。在地球上发现了超过5000种合成生物体自身物质的化学反应,可能的基因型一共有2的5000次方种(开与关),这个数字超过10的1500次方,也就是1后面跟着1500个0。2、蛋白质基因网络,生物利用20种氨基酸组成的蛋白质,构筑自身外形的控制基因。20种可能的氨基酸,将其比作字母,两个字母可以构成的文本就有400种(20的2次方),3个字母可以构成的文本就有8000(20的3次方)种,4个字母可以构成16万种。大多数蛋白质包含的文本长得多,即使是100个氨基酸构成的蛋白质,文本数量也超过了10的130次方种,即1后面有130个0。而人体的肌联蛋白,含3万个氨基酸。3、调控基因,生物调控代谢及蛋白质,形成各种性状的控制基因。通过这三大基因型网络,生命犹如积木,可以组合成近乎无数的生命形态,从而形成了地球这个多姿多彩的生命世界。[1]
不过,对于进化论的研究,仍有一些需确定和释疑的地方。比如,从概念来讲,“进化”一词明显包含了进步的意义。作者查阅权威的《现代汉语词典》,进化的涵义为:“事物由简单到复杂,由低级到高级逐渐发展变化。”[2]然而,这个定义与我们所了解的一些生物学事实相悖,并且已被不少严肃的生物学学者所批驳。举例来说,藤壶是一种附着于海边岩石上,有石灰质硬外壳的节肢动物。它演化出了一种粘附在螃蟹的下腹部,没有一定形状的寄生虫后代。这些后代以许多“根”攀住宿主螃蟹,其结构比藤壶简单,在水中摆动足部觅食。[3]另外,在一个哺乳类动物系统研究中选出的,24种祖先和后代有明显复杂性改变的物种,13组趋向简单,9组趋向复杂。[4]因此,生命并无复杂性的确定性趋向。同时,生命的高等和低等,也是个需要商榷的概念。例如美国权威刊物《科学(Science)》1993年7月发表的《免疫系统进化史》指出,低等生物的免疫系统,其复杂程度丝毫不亚于人类。[5]人体需要的13种维生素,我们自身只能合成两种,维生素d和b7,而大肠杆菌可以合成所有这些维生素;对于组成蛋白质的20种氨基酸,我们只能合成其中12种,而大肠杆菌可以合成所有这些氨基酸。[6]这样看来,连大肠杆菌都似乎比我们“高级”。还有一种看法是,进化会让有机体向分化的方向发展,但许多物种的化石证明,这也是种片面和错误的观点。比如马科曾经在远古有16种物种之多,现在仅存三种。[7]因此,一些生物学家认为,人们普遍认为的,进化过程是一个有机体向复杂、高级、分化的方向发展,进化表现一定的趋势,这些观点都是错误的。这个观点还认为,人类是“进化顶点”。然而,人类的产生,很有可能纯粹是机缘的巧合,这点我们将在未来讨论。生物遗传错误产生的变异是随机而漫无目标的,由于自然选择的缘故,而随着地域环境留存。因此,生命演进的历史,应当视为生命体系内部差异的增加和收缩,而没有趋势向的方向。复杂性的增加、物种的分化只是意外的结果。因此,为避免歧义,本文将避免使用“进化”一词,而采用更无趋向性的“演化”来描述生命的历史。
还有一个重要事实是,所有动植物,都是各自细胞与微生物紧密联合的共生体。每个动物身上分布着不同的生态系统:皮肤、嘴、肠胃、生殖器,以及任何与外界相连的器官——各处都分布着独特的微生物群,就像地球表面有各种各样的生态系统:雨林、草原、珊瑚礁、沙漠、盐碱地,每一种系统内都分布着不同的生物种群一样。以人体为例,人体内大约有30万亿个细胞,微生物的数量大约为39万亿。两者数量相差不大。然而,考虑到人体细胞分为有核细胞和无核细胞,有核细胞在人体中的占比约为十分之一。无核细胞没有细胞核,因此不具有复制自身的能力,他们主要参与人体内一些功能性的事物,比如红细胞用于输送氧气、血小板用于止血。无核细胞不是正常的有复制能力的细胞。其他微生物虽然有些是无细胞核的细菌等,但他们是正常的有复制能力的细胞。因此,如果将人体共生微生物和人体内可正常复制细胞数量进行对比,则比例达到了惊人的13:1。考虑到古代的卫生状况,古人的这个比例恐怕还要高上一点。甚至这还没完,每个动植物细胞,都有可能是一些小型生命的集群。动植物细胞中的线粒体、叶绿体、质体等细胞器,都有它们自己的DNA(构成基因的遗传物质)。也就是说,在很久远的过去,它们都是独立的生命个体。动物、植物、真菌等都属于真核生物。所有真核生物细胞都拥有同样的基本结构,即细胞dna包裹在细胞核内,因此称之为“真核”。细胞内部拥有像豆粒一样的线粒体,它们的工作是为细胞提供能量。真核生物可以追溯自20多亿年前,一个细菌在偶然情况下被另一个古菌(一种没有细胞核,但代谢途径又与真核生物类似的单细胞微生物,分布广泛,是地球生命的重要组成部分,是生物基本分类三域系统中的一域,另外两域为细菌域和真核域)所吞并,然而古菌没有消化掉细菌,两个生命体共生到一起,古菌提供了真核细胞的基本架构,细菌则最终转变成了线粒体。真核细胞生命演化出来后,它们中的一些开始合作聚集,最终产生了多细胞生物,乃至植物、动物等等。也就是说,真核生命起源于微生物的共生行为,也是一群微观生命的共生集合体。[8]
世界上千奇百怪的生命由此而来。如前所述,生命的基因变异需要经环境选择而留存。因此生物学中拥有一个重要概念“适应性”,定义为:“生物为了适应生存环境的变化而发生的在解剖学、物理学和行为学等方面的改变”。[9]那除了适应环境的变异以外,是否还有因其他因素保留的变异?实际上许多基因变异导致的性状改变对生物群体影响不大,比如让生物高矮胖瘦一点,不至于影响到生物群体的生存,这些随机的基因变异可以称得上是“中性”的,也会在生物群体中保留下来。然而,一些明显不是“中性”基因变异导致的性状差异,又似乎明显不利于物种生存,但却被大自然保存了下来。进化论的奠基人达尔文先生就发现,孔雀美丽的尾羽,对于生存明显是个累赘,已经影响到了飞行和奔跑,唯一的用处就在于吸引异性。因此,达尔文提出,动物中的雌性在选择配偶的时候,不只会考虑雄性是否有足够的“适应性”,还会选择那些看上去更美的配偶。雌性的审美偏好导致了雄性某些特别引人注目的特征,虽然这些特征不利生存,但仍然会保留下来。这就是性择理论。

孔雀美丽但略显累赘的尾羽[10]
性择理论提出后,学术界经过了非常长时间的争论,后续的理论主要分成了两派。一派认为性选择应归于自然选择,雄性华丽的特征是自然选择的强者所产生的盈余。对异性美的主观经验,很有可能是生物对适应度的潜意识评估。雄性被选择出来的某种特征,正反映了该雄性的实力,如尾羽漂亮庞大的孔雀,能够生存下来,一定孔武有力,不然就会被天敌猎食。这一流派的观点被不少实验所证实。比如,一个由科学家Craig Roberts领导的英国研究团队,从97个男性身上取出血液样本,测量主要组织相容性复合体(MHC)的三个基因的杂合度。MHC能使免疫系统分辨出自身细胞及外来病菌细胞,MHC杂合度越高,人体可能就越健康。研究人员在相同的灯光下拍摄这些男性的面部照片,然后将这些照片以随机顺序呈现给50位女性,由她们根据这些男性的吸引力高低来打分。结果发现吸引力越大的男性MHC杂合度越高。这就像这些女性只看男人脸部,便为他们做了一次精细健康检查。[11]另一派则认为,雌性的审美偏好与雄性炫耀特征之间是一个协同进化的进程。如果当初雄性的某种性状被雌性选中(如孔雀的尾羽),经过多代的群体选择,就会正向增强这种特征。群体中的雌性偏好这种特征的雄性,是因为她们觉得和这样的雄性能够生出一个漂亮孩子来,和他父亲一样,这个后代肯定也能够获得更多异性的青睐,从而延续自身的基因。最后这种性状就会变得一发不可收拾,使其异常出众。审美部分是生物独立于环境、自主的演化选择行为。这一派也提出了一些不可忽视的证据。在厄瓜多尔境内的安第斯山脉西部丛林中,生活着一种名为梅花翅娇鹟的小鸟。雄性梅花翅娇鹟在求偶时,会以每秒100次的速度快速振动和摩擦翅膀,发出高达1500赫兹的声音。这些声音响亮悦耳,非常独特,而且有真正的频率、音高、音调,很像小提琴的声音或者电话的拨号音。科学家们发现,雄性梅花翅娇鹟有独特的骨骼结构,尺骨性状与普通鸟类有较大不同,而且是实心的,其中的钙要比其他同科鸟类的同样骨骼多两三倍。地球上其他所有鸟类的尺骨,乃至雌性梅花翅娇鹟都是中空的。这种更宽更坚硬的实心尺骨有两个功能:一是通过为羽毛根部提供更坚固、稳定的锚定物来增强摩擦发声的效果;二是增强振动发声飞羽间的共鸣和耦合。这种改变明显降低了雄鸟的飞行能力。雌性梅花翅娇鹟的尺骨虽然也是中空的,但其形态也有同样的改变,因此它们的飞行能力也降低了,但付出的代价比雄性小。考虑到其他所有鸟类都有相同的柱状管翼骨结构,明显是在自然选择中实现最佳飞行能力的结果。然而梅花翅娇鹟由于审美偏好和炫耀特征,改变了翼骨结构,降低了飞行能力,这明显是与“适应性”以及个体实力无关的。
(左)雄性白冠娇鹟(梅花翅娇鹟同科鸟类之一)、(中)雄性梅花翅娇鹟和(右)雌性梅花翅娇鹟尺骨的X射线断层扫描图。图中的比例尺为2毫米[12]
这两派的讨论仍然没有明确的结果。未来如果有机会,作者也会将自己的一些相关思考与大家分享。
讨论了这么多生物学的内容,很多朋友可能会觉得,这些内容与本书的主题似乎无关。然而我们把眼光拓宽来看,人类和其他生物在地中海的演化过程,也是生物演化历程的一部分。如果我们无法理解这些基本事实以及生物演化的最重要力量,我们如何理解地中海的历史呢?所以之前的讨论绝对是有必要的,希望朋友们理解。下面,我们来正式进入地中海的生命世界。
在漫长的地球历史中,大多数生命因板块、地壳运动以及气候变化而分布和演化,它们随着地球的脉动生生不息。生命依赖于所处的环境,整个历程就像一场冒险,际遇或许是阻力,又或许是动力,有很大的运气成分。如前所述,533万年前地中海的形成毁灭了当时在盆地中居住的动植物,然而也为许多大西洋的生命们提供了一个新家。海豚、抹香鲸等海洋哺乳动物,常常通过直布罗陀海峡,游进地中海做客。

公元前1700-1300年 克里特克诺索斯 海豚及其他鱼类壁画。 克里特伊拉克列翁考古博物馆
海龟、海豹则是这里的常驻居民,并演化出了地中海僧海豹等本地物种。有些鱼儿也是地中海的匆匆访客。金枪鱼们发现这片平静的海域适合产卵,便年年来此产卵,之后洄游大西洋。人类在一万年以前(应该更早)便懂得捕捞这些地中

10000年前描绘金枪鱼的岩洞壁画
海定期旅客。近十几年来,由于金枪鱼是寿司的重要原料,地中海金枪鱼捕捞更是形成了年捕捞量五十万吨的庞大产业,并且影响到了金枪鱼的种群繁衍及相关生态系统。[13]随着时间的推移,常驻在地中海的海洋生物们也日渐繁多。不过如我们所知,地中海一直以蔚蓝清澈的海水知名,但科学家们发现,清澈也即水中的浮游生物和微生物较少,阳光容易到达较深海域。而浮游生物和微生物又是小型海洋生物的主要食物。地中海是一个低营养海域,有限的食物限制了海洋生物的生存繁衍。[14]接下来的冰期,剧烈的气候变动对海洋动物们影响巨大。科学家们发现,在300万到200万年前(我们可以回忆下那张气温变动图),冰川大量形成,锁住了巨量水分,导致海平面不断下降,海洋近岸区域消失,使得海洋动物经历了一次大灭绝事件。多达55%的海洋哺乳动物、43%的海龟、35%的海鸟和9%的鲨鱼物种,约三分之一的海洋大型动物在这次灭绝事件中消失。不过毁灭过后,新的生命形式如北极熊、海燕、黄眼企鹅等,大约四分之一的海洋动物物种逐渐出现,填补了空缺。[15]
在地面上,地中海周边地区受地中海对气候的影响,季节性变化显著。不少地区冬天湿润,少有霜雪;夏天炎热干燥,缺乏雨水,形成了一种独特的“地中海气候”。在这种情况下,许多植物都是在秋季和冬季生长,在雨水充足的春
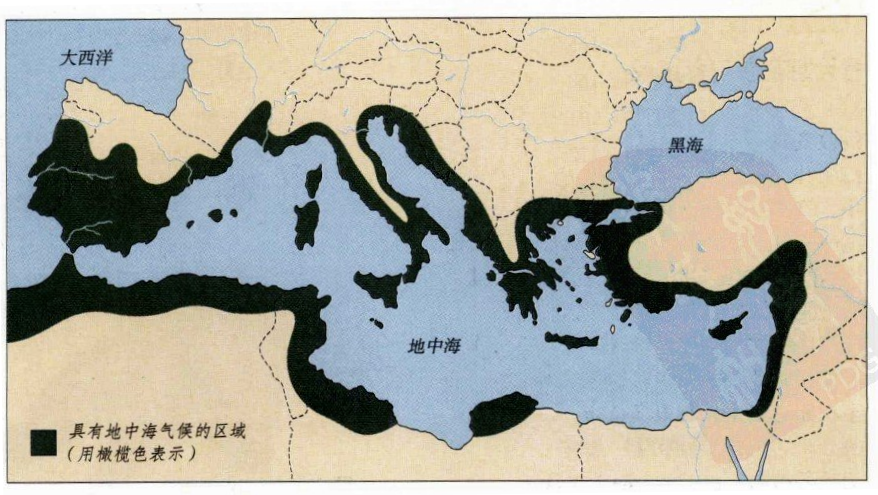
地中海气候区域图
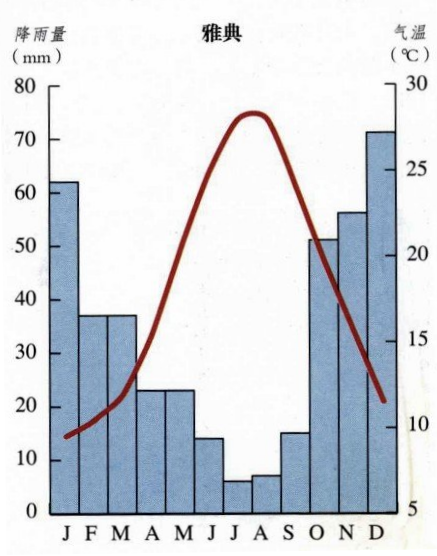
典型的地中海气候城市——雅典一年的气温和降雨量,红线代表气温变化,蓝线代表降雨量变化[16]
天开花。而在干热的夏天,绝大多数植物处于休眠状态,甚至有些植物会死亡。这也是起源于地中海的小麦主要是秋种春或夏收的原因。为了应对酷热的夏天,许多植物发展出了多样的生存技能。鼠尾草会在夏天将冬天的长叶枯落,取而代之的是卷曲、气孔更少的小叶子,同时分泌出一种香油覆盖在叶子表面,以降低水分蒸发,减少山羊的啃食(山羊不


喜欢这种香油的味道)。山柑在夏天仍然保持旺盛的叶子,不过那是因为它有很大的吸水根系;它甚至能在这时候开花,但只在凉爽的夜晚长出花朵,清晨浓郁的花香引来蜜蜂、蛾子等昆虫帮助其传粉;在白天酷热的太阳下,花朵非常容


易枯萎死去,花期仅有短短的一天。[17]不过我们不应当认为,地中海世界是非常严酷的生存环境。一方面,由于地中海的面积广大,海水吸收储存了大量的太阳热量。相对海洋来说,陆地表面吸收储存太阳热量能力差,因此白天热得快,夜晚凉得快;夏天热得快,冬天凉得快。当陆地温度较低时,地中海就向周边陆地输送热量,使得地中海地区在一天乃至一年的时间内,温度变化不至于过大。还有,地中海蒸发的水汽是这个地区水资源的主要来源,由于其面积较大,所带来的水蒸气也较多,整个地区的降雨量也大大高于内陆地区。最后,地中海世界并不都是地中海气候,周边地区气候非常多样。地中海就像空调一样,持续调节着整个地区的温度和湿度,使得地中海世界拥有非常宜居宜人的环境。大约有1万多种植物在这里生息繁衍,其中不少在后来被人类所利用,成为人类的重要资源。[18]
同样,冰期也对陆上动物产生了巨大影响。如前所述,在冰期低温时期,连欧洲南部都变成了冻土。冬天过去后,昆虫、蛙类等小型动物在这里又恢复了生机,成为鸟类觅食和繁殖的天堂。许多非洲的鸟类为了生存繁衍,开始了短途的迁徙。当时撒哈拉沙漠并没有那么大,从非洲草原到欧洲南部路程并不遥远。随着气候变暖,冻土逐渐往北消退,觅食地也随之北移,加上撒哈拉沙漠后来的扩大。如今,许多非洲的鸟儿需要横跨撒哈拉沙漠和地中海,艰辛地迁徙到繁殖区域。由于沙漠几乎没有食物和水,许多鸟儿在途中不幸死去。在冰期剧烈的气候变动情况下,鸟类的迁徙路线
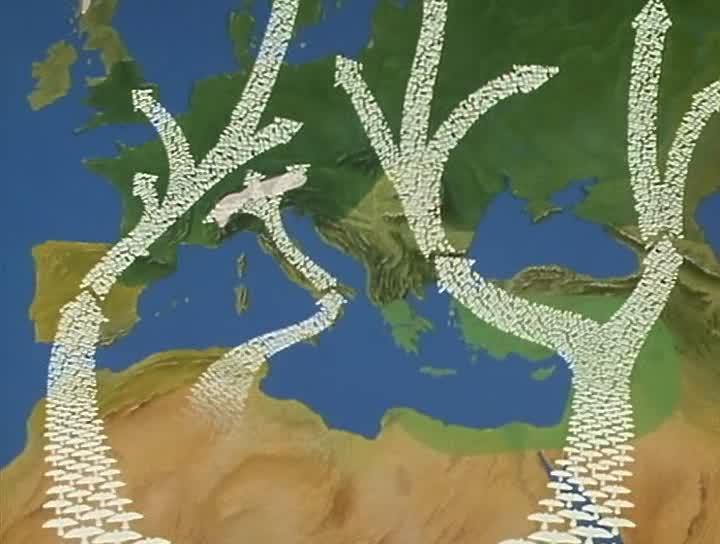
地中海周边鸟类迁徙路线示意图[19]
也随着冻土范围和撒哈拉荒漠的面积而延长和缩短。同样,亚洲南部的一些鸟类,也有在北方回暖后向北方迁徙的习性。这都是生物为了生存繁衍作出的努力。近三千年前的荷马曾在其诗中写道:“
如同多得无法胜数的群群飞禽,
有鸿雁、白鹤或脖颈修长的天鹅,
在亚细亚草原上,卡宇里奥斯河边
飞向东,飞向西,欣赏它们的翅膀的力量,
飞行后停下来大声啼鸣,使大地回响;”[20]
描写的正是这几种迁徙鸟类的行为。一些鸟类也有可能在留居在某地不再迁徙。如一些黑顶林莺已不再从欧洲的繁殖地迁往非洲。由于不需要长途飞行,这些林莺与它们迁徙的同类相比,翅膀更小。这些不迁徙的林莺,可能在未来分化出另一个物种。[21]
地中海诞生至今的时间正是哺乳动物飞速演化的阶段。如果我们能乘坐时光机器回到500万年前的地球,我们会发现除了非洲的犀牛类,找不到任何一种现存的哺乳动物。可以说哺乳动物的演化在漫长的地质年代中,周期显得极为短暂。由于哺乳动物种类众多,我们只能略微谈谈几类在当时常见的哺乳动物。
首先是有蹄类。有蹄类是我们熟悉的哺乳动物类群,像马、牛、猪、羊、鹿等。最早的哺乳动物都有5趾。有蹄的哺乳动物,由于植食性的自然选择,它们彻底改变并退化了趾头的数量、性状和构成。随着时间推移,它们的承重脚趾变大,形成蹄。这个特征持续演化了几百万年。从趾到蹄的持续演化令它们的脚变得强壮,从而增强其作为植食动物的奔跑等其他能力。有蹄类通常有强壮的臼(jiù)齿(即研磨和咀嚼食物的磨牙),用以咀嚼植物粗糙的茎叶。为了消化植物粗纤维,有蹄的哺乳动物演化出了多室肠道,这些肠室寄生着多种共生微生物,帮助它们把植物细胞壁的纤维素和木质素转化成可消化的营养。通过这些演化,有蹄类成为陆地上拥有主导地位的植食性动物。牛、鹿等动物还通过反刍(chú)来增强自身对植物的消化能力。它们对嘴里的食物稍加咀嚼,就吞咽下去。食物下到第一个胃室后,在那里被部分消化,然后返送回嘴里,进行二次咀嚼然后真正吞咽下去。有蹄类种类众多,并且有多种起源。甚至鲸类(鲸鱼和海豚)在分类学也上属于有蹄类,他们一开始是陆上的有蹄类哺乳动物,随后退回到了海洋环境中。近期化石证据和分子分析综合研究表明,鲸鱼最近的陆地亲戚有可能是河马,与牛、猪等有蹄类的关系次之。以下是500万年来,我们在地中海周边常见的几种代表性有蹄类动物:
1.鲨齿鲸(学名Squalodon sp.):生活于渐新世晚期到上新世时期,全长2-2.5米。鲨齿鲸眼眶上有个突起肌肉,它们可能通过这个肌肉和鼻子上的器官,发出超声波来进行定位。它们在外表上与海豚相差无几,但牙齿保留了古鲸类的原始特征。

2.三趾马(学名Hipparion sp.):在马、牛、猪这些有蹄类动物中,马拥有独立的起源。马、蝙蝠和食肉类哺乳动物的共同祖先首先从其他哺乳类中独立出来,之后,马独自演变为有蹄类动物。因此,马虽然从演化历史上与牛、猪等没有亲缘关系,但由于相同的环境及习性,演化出相同的适应性成为有蹄类。三趾马生活于中新世中期到更新世,在亚欧非乃至北美洲都有广大分布。肩高约1.5米(肩高指肩部到脚掌与地面接触面的垂直距离,可以理解为动物低下头的最大高度)。和现在的马相比,三趾马还保留有三根趾,但两侧的趾已经退化到够不到地面了,只有增大了的中间趾(蹄)具有承重功能。随着新生代以来的气温降低,许多森林变为草原或半草原状态。因此三趾马的祖先从多趾趾行逐步演化为蹄行。同时体型逐渐增大,牙齿演化得更加适应嚼食草茎。臼齿带凸起尖脊,可更加高效地碾磨植物。它们的消化系统也变得有利于消化植物中粗糙的多纤维碳水化合物。这些演化是它们适应草原自然环境的表现。

3.披毛犀(学名Coelodonta antiquitatis)和板齿犀(学名Elasmotherium sp.):犀牛的角不是骨质,而是主要由与毛发同样成分的角蛋白构成的,因此一般不会保留为化石。但长角处的头骨表面会变得很粗糙,由此可知角的存在。披毛犀生活于上新世晚期到更新世末期的亚欧大陆,是适应冰期寒冷气候的食草动物,身长可达4米。板齿犀则生活在更新世早期到中期,身长更可达4.5米,肩高可以达到2米。一个很特别的是,板齿犀的门齿已经消失,它们是用嘴唇来拔草食用。


4.果猪(学名Metridiochoerus sp.)和南猪(学名Notochoerus sp.):这两种当时非洲猪类生物的代表都是长有长獠牙的杂食动物。身长可达2-2.5米。果猪生存于上新世晚期到更新世中期,臼齿齿冠(即露出牙龈的牙齿部分)高,据此推测其更适应粗纤维食物;南猪生活于上新世早期到更新世早期,齿冠不是很高,因此可能更适应细纤维食物。

5.西瓦兽(学名Sivatherium sp.),与长颈鹿有可能的亲缘关系,因此被归于长颈鹿科。在上新世早期-更新世晚期的亚欧非大陆有广泛分布。肩高可达2-2.2米,头骨长度可达0.7米。撒哈拉沙漠的岩画遗址也有类似西瓦兽的生物,因此它们可能一直生存到全新世。

6.朱马麟(学名Giraffa jumae),生活于上新世早期到更新世晚期的非洲,全高可达5米。可以理解为长颈鹿古代种。
7.枝角鹿(学名Eucladoceros sp.)和大角鹿(学名Megaloceros giganteus)。鹿科是有蹄类一个非常繁盛的类群。起源于中新世早期甚至更早,以欧亚大陆的温带-寒带森林为中心向外辐射,分布广泛。鹿角每年都会替换,一般只有雄鹿才有,只有驯鹿才是雌性也有角。枝角鹿和大角鹿都分布于亚欧大陆,肩高约1.8-2米。枝角鹿生活于上新世晚期到更新世中期,大角鹿则生活于更新世晚期。枝角鹿的角左右宽度最大可达1.7米。大角鹿的角更可达3米多宽,总重可达45公斤,是拥有已知最大角的鹿科动物。为了支撑巨角,其脖子到肩膀的脊椎和肌肉非常发达。在受到人类捕猎后,大角鹿种群逐渐萎缩,最后灭绝。不过近期的化石发现,距今7000年前,在中亚的乌拉尔山脉,这个物种还存在着。

8.草原野牛(学名Bison priscus):生活于更新世中后期的欧亚大陆及北美洲,肩高可达2米的牛科动物。是牛科中最大的种类之一,体重可达2吨。

9.原始牛(又名原牛,学名Bos primigenius):生活于更新世晚期到全新世的欧亚大陆,肩高约1.8米。是家畜牛的祖先。角两端之间的宽度最大可达1.5米,角细长。有可能于1627年灭绝。

10.欧式惊奇巨羊(学名Pelorovis oldowayensis):虽然称之为巨羊,但仍然是牛科的一种,生活于更新世早期的非洲,肩高约1.5米。头骨长,主要食草。角的长度可达1米,并且在演化过程中也形成了多种形状。

11.以撒巨羚(学名Megalotragus isaaci),狷羚类生物。生活于上新世晚期到更新世早期的非洲,肩高约1.4米,鼻子额头高耸。

12.妖眼河马(学名Hippopotamus gorgops),生活于更新世非洲的河马类生物。身长可达4-5米。眼睛的位置比现代河马高。大足便于在河泥上行走,獠牙是为了挖掘水草。和现代河马一样,喜欢呆水中,四肢强壮,主食水生植物。

接着是大象和它们的亲戚,被称为近蹄类,意思是“差不多就是有蹄类”。大象可能起源于始新世的北非,祖先形体大小如猪,可能生活在淡水湖泊和池塘附近。在那里它们完成了大部分早期进化。大象的化石遗骸很丰富,这归因于它们的体型较大,令他们的遗骸比小型动物容易保存得多。大象的种类曾经多种多样,但现存仅有分布于亚洲和非洲的3个物种。近蹄类的共同特征是增大的头骨,外显的象鼻,粗短的脖子,支撑庞大身躯的扁平足柱形长腿。大象颌部的第二对上切齿变长,延伸为象牙。象牙可能刚开始用来收集植物,随后演化成一种展示自我的装饰。随着演化,大象的臼齿变得更深更宽,有更多的脊,形成明显的脊齿型研磨面,适合研磨植食。
1.恐象(学名Deinotherium sp.):恐象生活于中新世到更新世中期的亚欧非大陆,分布广泛。它是最大的近蹄类动物之一,肩高3.5米,比现代大象平均高出1米。与其他大象向前的象牙不同,它有一对锋利的朝下的象牙,在下巴下方向后弯曲。对于恐象象牙的用途,有多种猜测,可能用于连根拔起植物,拖拽树枝,以及撕剥树皮等。

2.真猛犸象(学名Mammuthus primigenius),更新世中晚期生活于亚、欧、美洲的北部。真猛犸象是适应冰期寒冷气候的象类,全身披长毛,皮下脂肪厚,耳朵小,象牙大幅度弯曲,肩高3-3.5米。猛犸象臼齿精巧,高而宽,深脊,有珐琅覆盖,非常适合研磨坚韧的干枯植物,比如长在北半球寒冷的干燥草原和林地的草、灌木等。最早的猛犸象没有毛发,大约700000年前,当冰期到来,猛犸才演化成有毛动物。末次冰期后,随着人类过度猎杀和栖息地变暖和可能的疾病传播等因素,猛犸象种群大幅萎缩。最后一只猛犸象可能在3500多年前,于北冰洋的朗格岛死去。

3.互棱齿象(学名Anancus sp.),上新世到更新世生存于欧洲的象类,是猛犸象的一个欧洲亲戚。它的象牙约等长于其身高,最高可达3米。其牙齿最适合咀嚼云杉叶和其他柔软植物,这一点与猛犸象适合进食坚韧粗糙的植物不同。

食肉类。大多数现存的食肉哺乳动物都是食肉类成员。现代食肉类祖先是古新世早期的食虫动物。在很早就分裂成两大类群:猫型亚目(猫、麝猫和土狼)犬形亚目(狗、鼬鼠和熊)。两者区别主要是犬形亚目颌部后方有发达的大型粉碎性臼齿,适应碾碎骨头,而猫型亚目没有这样的臼齿。猫型亚目和犬形亚目的另一个区别在于内耳的骨结构,猫型亚目类的内耳包含在软骨内,而犬形亚目的内耳包含在骨头内。现存的食肉类动物是这些宗亲的现存代表。除了陆地的猫型亚目和犬形亚目,食肉类的第三个演化支系是水生的鳍脚目,包括海豹、海狮、海象等。这些海洋动物是从犬型亚科的熊这一支进化而来。把食肉类统一起来的主要特征是它们的牙齿,所有的食肉类,颌部每一侧都有一对裂齿,每一对裂齿都有与其合作的增大上下臼齿,颌部闭合时,这些牙齿就成了两对切肉剪刀。绝大多数食肉类都有这种牙齿,一些特别的动物,如海豹,由于适应容易咬开的鱼类,裂齿就消失了。不少食肉类还有用来夹紧食物的锋利切齿,以及用来攻击的犬齿。
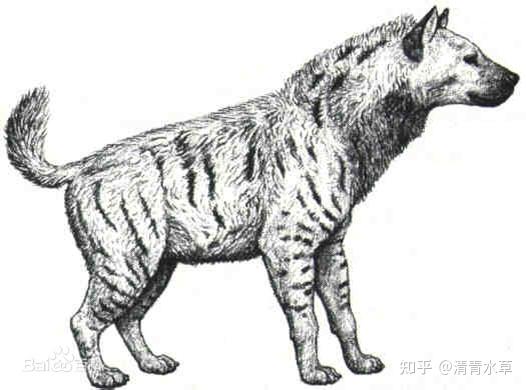
1.硕鬣狗(学名Pachycrocuta),上新世晚期出现,更新世中期在欧亚非广泛分布。鬣狗类动物以强力咬合著称,前腿比后腿长,擅于奔跑,可耐心地跟踪猎物好几公里。最早的鬣狗类动物是中新世早期的小型动物,可能主要食虫,与麝猫类似。硕鬣狗可能是最大型鬣狗类动物,站立肩高1米,狮子大小,体格结实粗壮。不过与现在的鬣狗不同,较短下肢令其不擅奔跑和长距离追猎,更擅长力量型近距离攻击。为了食物,它也会和大型猫周旋,偷取它们的猎物尸体,甚至可能食腐。在末次冰期之后,硕鬣狗开始灭绝,灭绝时间几乎与剑齿猫同步。可以预料剑齿猫的消亡原因与巨型鬣狗的灭绝有关。而那些速度更快的鬣狗群体存活了下来。不过它们的分布地域逐渐缩小,当前仅局限于非洲和热带印度地区。
2.更新世洞熊(又名熊鼠,学名Ursus spelaeus),更新世分布于欧洲到中亚区域,是现代灰熊的祖先,冬天可在洞穴冬眠。最早的熊在中新世演化出来,熊的牙齿适应各种食物,食谱限制少。依靠这项技能,中新世晚期到更新世的成功熊类郊熊,已在亚欧非大陆广泛分布。郊熊的腿比洞熊长,肩高也高于洞熊。洞熊在末次冰期广泛扩散,颌部强健,颊齿大而平,可以研磨坚韧的植物乃至骨头。在欧洲的某个洞穴发现了数以千计的洞熊样本,应该是冰期寒冬,巨熊聚集在洞穴的体现。洞熊身长约2米,体形巨大;头骨大而长,有些长度超过了0.5米;不过腿较短。洞熊曾与早期人类共存,并被其所追猎。

3.短剑剑齿虎(学名Machairodus sp.),在中新世晚期到更新世早期广泛分布于亚欧非,乃至北美。肩高约1.2米。在非洲是主要的化石猫类,剑齿猫类最大的种类。头部长度约0.35米。与现生狮子几乎相同,只是宽度窄细一些。

4.洞狮(学名Panthera selaea),更新世中后期广泛分布于欧亚大陆,身长2-2.7米,比现在的狮子要大,骨骼形态类似于现在的狮子,约1万年前灭绝。从古人类的洞穴岩画中可以看出,洞狮没有现代狮子类似的鬃毛,尾巴尖端也没有毛球。喜欢寒冷干燥的草原环境,以马、鹿、羚羊等为食。也有人认为它为现代狮子的祖先。

啮[niè]齿类动物。上下颌的门齿没有牙根,直接从颌骨生出,能终生生长,是啮齿类动物的共同特征。由于门齿不断生长,使它们需要不断啃啮木头、坚果和其他坚硬物质,以磨损门齿,避免太长后无法进食。现生物种超过1800个,种数约占哺乳动物的 40%~50%。啮齿总目有两个亚群,啮齿目和兔形目,由共同祖先演化而来。啮齿目上下颌各有一对门齿,最早成员长得更像松鼠而不是老鼠。古新世晚期,它们在北美洲首次辐射,逐渐扩散到各地的适应生态位。它们是一个特别成功的进化支,大鼠、小鼠和它们的宗亲成为最多样化,也可能是最普遍存在的现生哺乳动物类群。它们常见的成员还有生活在林地的松鼠,占据水域的海狸和河狸,南美洲的家畜豚鼠等。它们的颌部结构也很特殊,当他们啃啮时,颌部底部可以伸出,从后向前咀嚼。兔形目有家兔、野兔和鼠兔。与啮齿动物不同,它们有两对凿子似的上门齿,取代了啮齿目的一对。兔形目颌部构造也不同,它们的咀嚼采用横向咀嚼的方式。后肢很长,构造特别适合弹簧似的蹦跳。最早的兔子起源于古新世晚期到始新世早期的亚洲,是最成功的植食性动物之一。
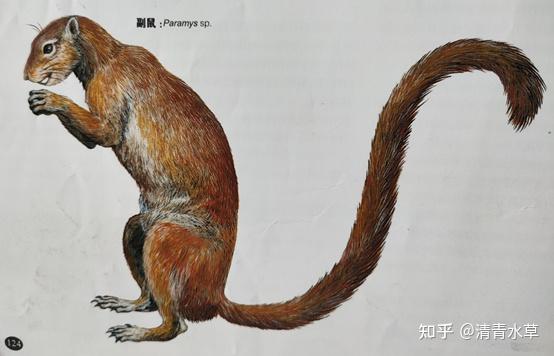
1.副鼠(学名Paramys sp.)副鼠在古新世末期到始新世中期生活在欧亚大陆及北美洲,不含尾巴的身长约45-60厘米。是各种特征都比较原始的啮齿类动物。
2.模鼠(学名Mimomys sp.),中新世晚期到更新世晚期生活于欧亚大陆,不含尾巴的身长约10厘米,可视为田鼠的古代种。
3.副古仓鼠(学名Paracricetodon sp.),渐新世生活在欧洲,不含尾巴的身长15-20厘米,是早期的仓鼠种类。

4.飞松鼠(学名Miopetaurista sp.),中新世早期到上新世生活于欧亚大陆和北美洲的温暖森林中。是大型飞鼠的一种。形态、生态与现存的飞鼠——大鼯鼠相似,可以在树木间滑翔飞行。

5.上新五褶兔(学名Pliopentalagus sp.),中新世后期到上新世生活于欧亚大陆,身长约0.4米。

最后,是灵长类动物。今天,在美洲、非洲和亚洲生活着超过200种非人类灵长类动物。灵长类和其他哺乳动物的区别在于以下这些特征:
- 可灵巧抓握的手指和脚趾,因而它们能轻松抓住树枝,在树上生活。灵长类有强烈的杂食性倾向,他们的臼齿齿冠(牙齿露出牙龈的部分)低,齿尖钝,适合消化各种各样的食物,有根据环境改变饮食习惯的能力。双眼视觉。许多动物的眼睛位于头骨两侧,如松鼠、鱼,视野不会重叠。灵长类的眼睛面向前方,双眼视野重叠,拥有立体视觉。对于在树枝蹦跳的动物来说,立体视觉可帮助判断树木间距,无疑极为重要。增大的大脑。相较于其他哺乳动物,灵长类的某些感觉有所退化,如听力嗅觉不如狗等。但灵长类在进化过程中,大脑的其他部分有所增大。更大的脑容量令智力和适应能力都大大加强。这是灵长类成功的关键因素。
除人类外灵长类动物多数栖息于气候稳定且食物充足的丛林地带,即使有迁徙行为也多是在林地内进行短距离迁徙。极少数也可生活在陆地上。
- 中猴(学名Mesopithecus sp.),中新世晚期到上新世生活于欧洲、伊朗等地,身长约0.6米,是现今疣猴类的直系祖先。雌雄个体差异大,身体骨骼、犬齿等在雌雄个体间有很大差别。现今疣猴类多数为树栖,但中猴也有可能为陆栖。欧氏狮尾狒(又译奥氏狮尾狒,学名Theropithecus oswaldi),是更新世生活于非洲的猴类,肩高约0.8米。在当时非洲分布广泛,个体众多。雌雄个体差异大,有雄性个体可达雌性大猩猩的体重。

[22]
- 以上综述来自《适者降临》作者:(美)安德烈亚斯·瓦格纳,译者:祝锦杰 浙江人民出版社2021年版;《生命的运作方式》作者:(美)马伦·霍格兰(Mahlon Hoagland) /伯特·窦德生(Bert Dodson),译者:洋洲/ 玉茗 后浪丨北京联合出版公司2021年版;《进化论与生活》作者:(美)戴维·斯隆·威尔逊,译者:李明芝,中国人民大学出版社,2021年版。 ↑ 《现代汉语词典》作者: 中国社会科学院语言研究所词典编辑室 编 出版社: 商务印书馆1996年版,第659页。原谅作者手头只有这么早的版本...... ↑ 《生命的壮阔——从柏拉图到达尔文》作者: (美)史蒂芬·杰·古尔德 译者: 范昱峰,江苏科学技术出版社2009年版第112页 ↑ 前引书 第169页 ↑ 前引书 第15-16页 ↑ 《适者降临》作者:(美)安德烈亚斯·瓦格纳,译者:祝锦杰 浙江人民出版社2021年版第065、078页 ↑ 《生命的壮阔——从柏拉图到达尔文》作者: (美)史蒂芬·杰·古尔德 译者: 范昱峰,江苏科学技术出版社2009年版第5章。另该书称马科只有一种物种,但马科马属下有马,驴和斑马,因此改为三种。 ↑ 《我包罗万象》 作者:[英] 埃德·扬 译者: 郑李 出版社: 后浪丨北京联合出版公司2021年版 1.生命的岛屿 ↑ 《史前地球》丛书中的《哺乳动物的时代》,作者:[美] 托姆·霍姆斯 译者: 邬冬文,上海科学技术文献出版社2021年版 第138页 ↑confused_me先生的摄影作品https://pixabay.com/zh/photos/peacock-bird-plumage-feather-2490176/ ↑ 可参见自然选择的另一位提出者华莱士Alfred Russel Wallace的观点及以色列生物学家扎哈维Amotz Zahavi 提出的“不利条件原理”。实验资料来自《进化论与生活》作者:(美)戴维·斯隆·威尔逊,译者:李明芝,中国人民大学出版社,2021年版,第74-75页 ↑ 可参见现代统计学的奠基人、生物学家罗纳德·费舍尔(Ronald Aylmer Fisher)提出的失控理论(Fisherian runaway),以及美国鸟类学家理查德·普鲁姆(Richard Prum)提出的审美假说。例子来自《美的进化》作者: [美]理查德·O.普鲁姆(Richard O. Prum)译者: 任烨 / 审校:刘阳,中信出版社2021年版,第4章 审美的创新与物种的衰落 ↑ 纪录片 《The First Eden》作者(英) 英国广播公司BBC,1987年;《超鱼(精编版)》: 央视网2021-11-04,http://tv.cctv.com/2021/11/04/VIDE1415031674825325.shtml(暂未找到原资料出处);《地中海金枪鱼捕捞战》,作者:维维恩·瓦尔特, 译者:关林,来自期刊:国外社会科学文摘, 2006, 000(011):38-39,原载于美国《时代》杂志2006年8月11日。 ↑ 纪录片《Oceans-07.Mediterranean.Sea》作者(英) 英国广播公司BBC,2008 ↑ 《科学大观园》 2021年第18期 P58页《远古时代的海洋动物大灭绝》作者:科东方,来自瑞士苏黎世大学和德国柏林自然博物馆的科学家研究发现 ↑ 图片引自《野花:地中海沿岸地区500多种野花的彩色图鉴》作者:(英)伯尼,译者:张扬,中国友谊出版公司2008年版 第6页 ↑ 纪录片 《The First Eden》作者(英) 英国广播公司BBC,1987年 主持:大卫·艾登堡 ↑ 数字引自《野花:地中海沿岸地区500多种野花的彩色图鉴》作者:(英)伯尼,译者:张扬,中国友谊出版公司2008年版 第6页 ↑ 纪录片 《The First Eden》作者(英) 英国广播公司BBC,1987年 主持:大卫·艾登堡 ↑ 《荷马史诗·伊利亚特》作者:(古希腊)荷马;译者:罗念生,王焕生;人民文学出版社,1994年版,第45页 ↑ 《大迁徙》作者:(英)霍尔;译者:平晓鸽;中国大百科全书出版社,2021年版第48-50页。 ↑ 以上对哺乳动物的讨论和图片综合来自:《进化伊甸园》作者:(美)阿兰·特纳,莫西奥·安东;译者:闾春晖;江苏科学技术出版社2021年版。《灭绝的哺乳动物图鉴》作者:(日)富田幸光,;译者:张颖奇;科学出版社2021年版。图片大多来自该书。《史前地球》丛书中的《哺乳动物的时代》,作者:[美] 托姆·霍姆斯 译者: 邬冬文,上海科学技术文献出版社2021年版。以及《生命世界》期刊哺乳动物相关文章。 ↑
2021年4月20日
[第二章、地中海世界的生命] 相关文章推荐:
- 最新经典文章
- 热门经典文章
- 热门文章标签
全站搜索